你有没有注意过这样一个现象:二十岁时饿一顿,第二天上秤能轻两斤;到了五六十岁,哪怕连着一两天吃的很少,肚子上的肉还是纹丝不动,甚至腰围还在涨。
很多人把这种现象归结为“代谢慢了”,但真正的原因比这复杂得多。

5月20日,哈佛大学公共卫生学院的研究团队发表在《Nature Aging》期刊上的一项研究指出,问题的根源可能不在我们常以为的线粒体身上,而是一个长期被忽视的细胞器——过氧化物酶体[1]。
(文章最后有归纳总结和现阶段的实用小建议!)
什么是“代谢灵活性”?——年轻时是混动,老了变单燃料
首先,我们要理解一个概念:代谢灵活性。简单说,就是身体在进食和饥饿之间切换时,能自如地选择燃烧糖还是脂肪。吃饱了,就优先用葡萄糖供能;饿了,则迅速切换到燃烧储存的脂肪。
年轻时,这套切换系统很灵敏。但上了年纪后,代谢灵活性会显著下降——身体变得越来越“固执”,只愿意烧糖,脂肪则堆在细胞里怎么都动不了[2]。这就能解释,为什么很多中老年人明明吃得不多,却依然有脂肪肝、腹部肥胖,甚至发展成代谢综合征。

过去几十年,科学界主要把目光放在线粒体上。线粒体是细胞的“发电厂”,它的功能随年龄下降,确实是衰老的重要标志。但问题是:线粒体出状况,究竟是衰老的原因,还是结果?
这项新研究给出了一个出人意料的答案:线粒体可能是被“拖累”的,真正的 “罪魁祸首”是上游的过氧化物酶体。
过氧化物酶体:细胞里的“危化品处理厂”
过氧化物酶体的体积很小,直径只有几百纳米,在细胞里存在感不强。但它干的都是“脏活累活”,专门分解那些线粒体处理不了的特殊脂肪,比如超长链脂肪酸(碳链长度≥22)和某些多不饱和脂肪酸(PUFA)[3]。
要完成这些任务,过氧化物酶体需要把几十种不同的酶“请”进自己身体内部。这个过程依赖一个关键蛋白,在线虫中叫PRX-5,在人类和小鼠中叫PEX5。你可以把它理解为一张“门禁卡”,没有它,那些分解脂肪的酶就进不去过氧化物酶体。
研究团队首先比较了年轻(1日龄)和年老(8日龄)线虫在禁食状态下的基因变化。他们发现,年轻线虫在禁食时会迅速启动一组特定基因,而这组基因里最富集的,就是过氧化物酶体相关的蛋白输入和脂肪酸氧化基因(比如prx-5、ech-8、daf-22)。但在年老线虫中,这种诱导反应几乎完全消失。
这意味着:随着年龄增长,细胞失去了在需要时“激活”过氧化物酶体的能力。
PRX-5水平下降,过氧化物酶体变成空壳
为了直接观察过氧化物酶体的功能变化,研究人员用荧光蛋白标记了进入过氧化物酶体的酶(GFP-SKL)。在年轻线虫的肠道里,荧光呈清晰的点状——说明酶被正常运了进去。但在10日龄的老年线虫中,这些点状信号大幅减少,荧光变得弥散,说明酶进不去了。
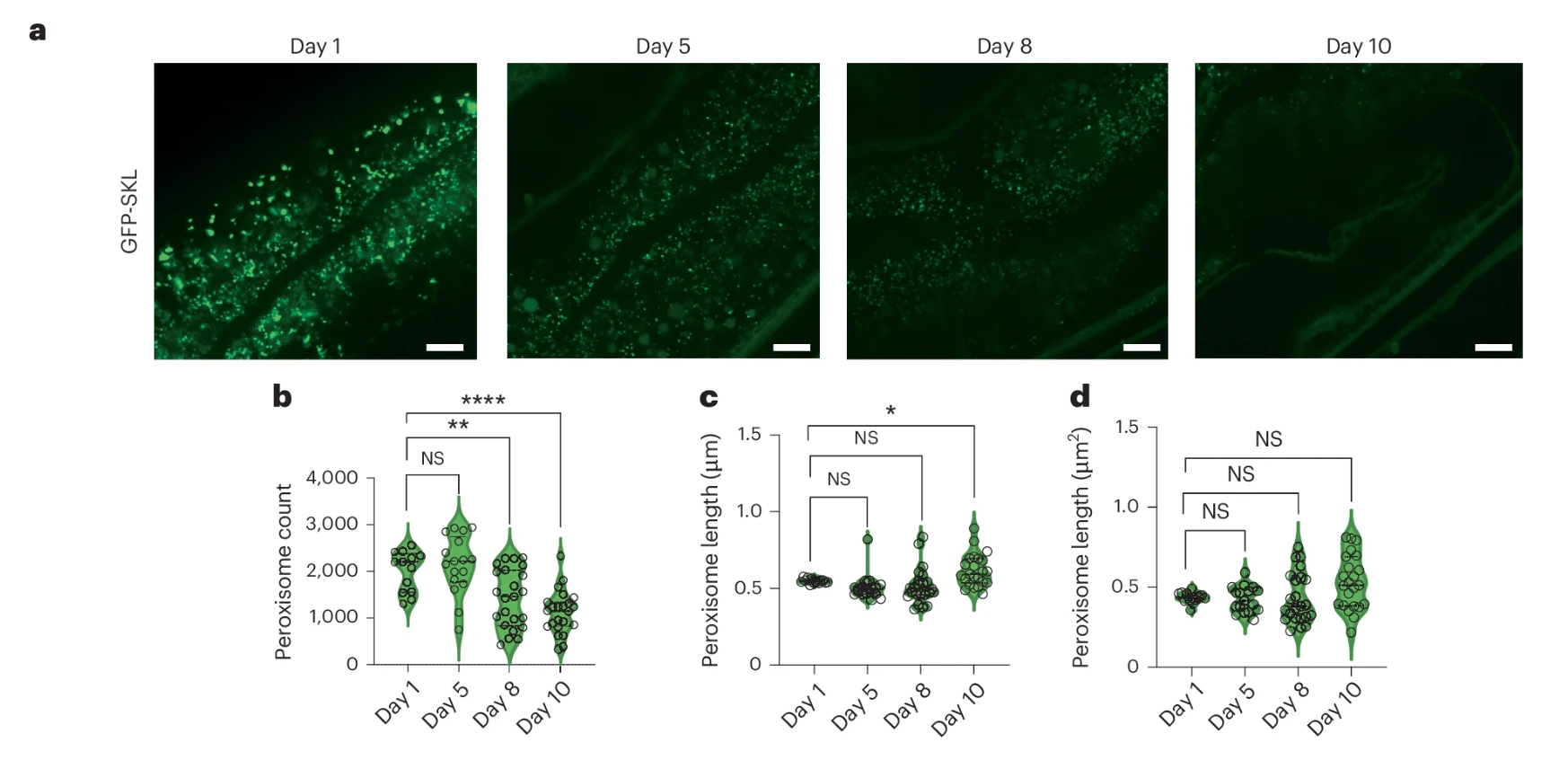
与此同时,PRX-5蛋白本身的水平也在急剧下降。定量分析显示,从1日龄到12日龄,PRX-5的量减少超一半。
这还不止。研究人员发现,随着年龄增长,过氧化物酶体开始与溶酶体融合,发生管状化并被降解。也就是说,不光是“门禁卡”丢了,连“垃圾处理厂”本身都在被细胞回收拆除。
脂肪“焊死”在细胞里,为什么饿也饿不动
接下来是最关键的一步:过氧化物酶体坏了,对代谢到底有什么影响?
研究团队构建了prx-5基因缺失的线虫。结果非常明显,这些线虫即使不进食,体内的脂滴也比正常线虫大得多,数量也多得多。更严重的是,当研究人员让这些线虫禁食12小时,正常线虫的脂肪储备显著减少,而prx-5缺失线虫的脂肪纹丝不动。
这就是典型的代谢僵硬,身体失去了动员脂肪的能力。
为了搞清楚哪些脂肪“卡住”了,团队做了脂质组学分析。他们比较了正常线虫和PRX-5缺失线虫在禁食前后各种脂质的变化。结果发现,正常情况下,禁食会消耗掉一大批含有EPA(二十碳五烯酸,一种ω-3多不饱和脂肪酸)的甘油三酯(TGs);但在PRX-5缺失的线虫中,这些EPA-TGs不但没减少,反而堆积在一起。
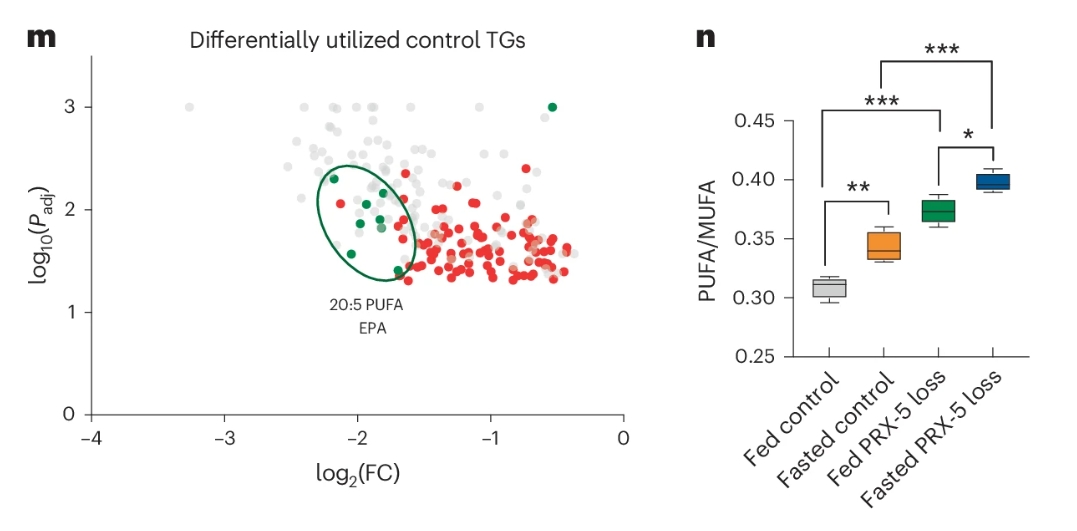
更麻烦的是,PRX-5缺失还会导致细胞内多不饱和脂肪酸与单不饱和脂肪酸的比例(PUFA/MUFA)显著升高。高比例的PUFA会促进脂滴之间的融合,形成体积更大的“超级脂滴”,而这些大脂滴更难被分解。
线粒体是怎样被“连累”的?
过氧化物酶体坏了,线粒体当然也逃不掉。
研究人员使用了一种诱导型降解系统(AID),可以在成年早期精准地“关掉”PRX-5。结果发现,在PRX-5被降解后的48小时内,线粒体就开始出现异常——肿胀、碎片化,原本网状的结构变得支离破碎。
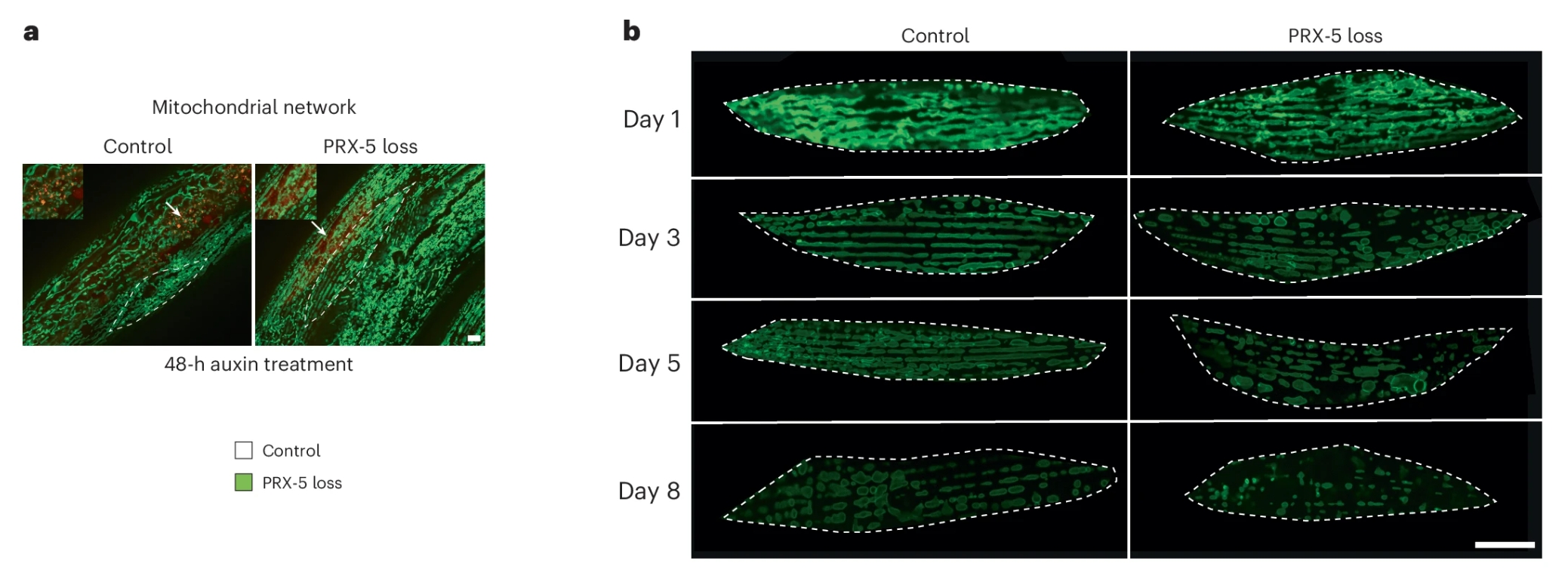
更值得注意的功能变化是线粒体膜电位。正常的线粒体膜电位维持在一个相对狭窄的范围(约-130到-180mV)。但在PRX-5缺失的情况下,线粒体膜电位出现了明显的超极化,也就是比正常更负。这听起来像是“更有活力”,但实际上是一种病态。长期的超极化会引发电荷失衡、活性氧增加,最终走向去极化和功能崩溃。
为了确认这些研究结果在哺乳动物细胞中也成立,研究团队使用了PEX5基因敲除的人源A549细胞。结果与线虫一致,PEX5缺失的细胞基础耗氧率和最大呼吸能力都显著下降,而糖酵解水平(反映细胞对葡萄糖的依赖)则明显升高。也就是说,没有功能性过氧化物酶体,细胞被迫放弃高效的氧化磷酸化,转而依赖低效的糖酵解来生产ATP。
这个时间顺序很关键:先有过氧化物酶体失灵,后有脂滴堆积和线粒体崩坏。 过去很多被认为是“线粒体衰老”的现象,其实可能是过氧化物酶体先出了问题。
转机:饮食限制和基因修复
那有没有办法逆转这个过程?
研究团队测试了饮食限制(DR,也就是“少吃点”)。他们把线虫放在低浓度细菌的培养基上,模拟长期热量限制。结果发现,年老(8日龄)的饮食限制线虫,其过氧化物酶体功能依然保持得相当好,与年轻线虫没有显著差异(图5b-c)。
但有一个更关键的实验:如果在饮食限制的同时,诱导PRX-5降解,那么饮食限制带来的寿命延长效果完全消失(图5d)。这说明,饮食限制之所以能延寿,在很大程度上有赖于它保护了过氧化物酶体的功能。
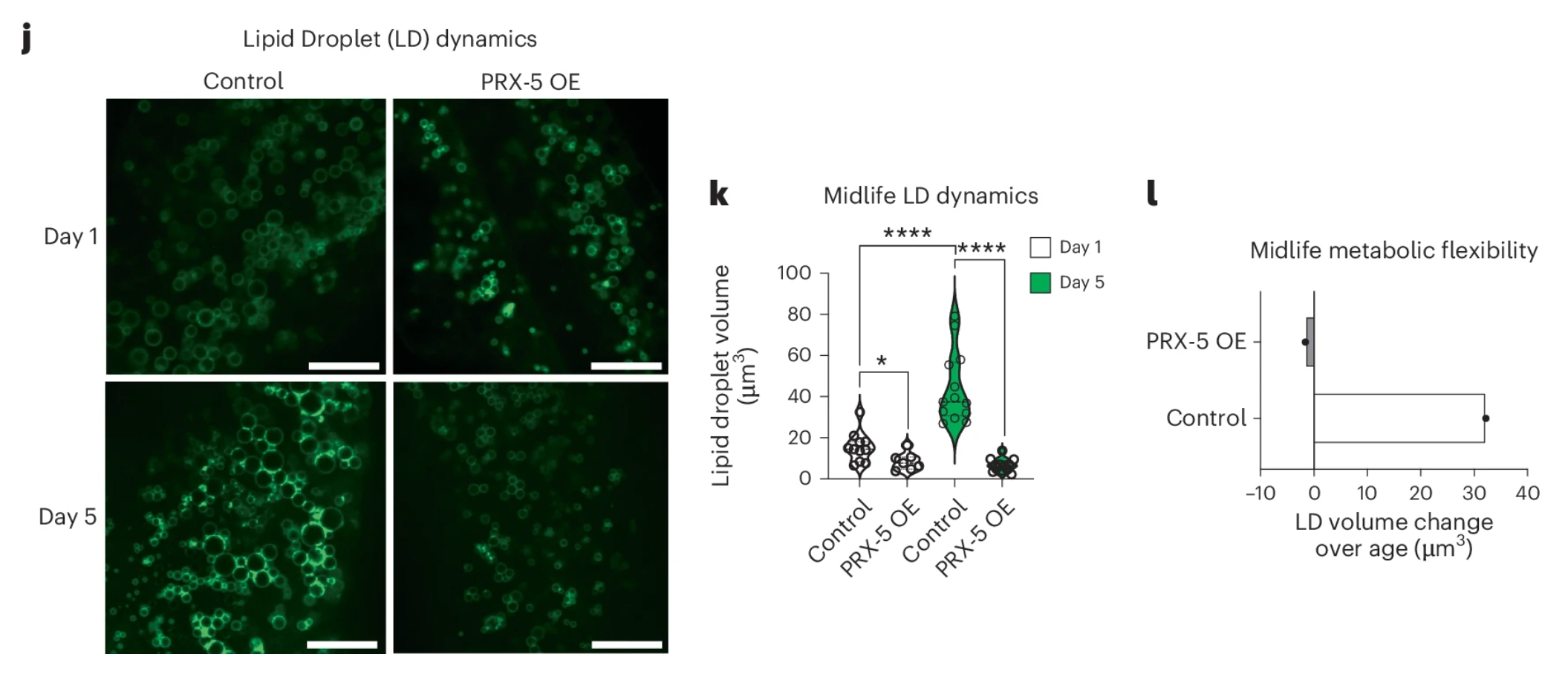
更直接的证据来自基因干预。研究团队构建了过表达prx-5的转基因线虫,这些线虫在中年(5日龄)时的脂滴大小显著小于对照组,老年(8日龄)时的线粒体形态也更健康(图6j-l)。更重要的是,prx-5过表达将线虫的中位寿命延长了约15-24%,最大寿命也显著提升。
那是什么在调控prx-5的表达呢?研究团队找到了上游的转录因子 NHR-49(人类中的同源物是PPAR-α,一种著名的脂肪酸氧化调节因子)。在饮食限制的线虫中,NHR-49更多地停留在细胞核里,处于激活状态,从而上调prx-5等过氧化物酶体基因的表达。随着年龄增长,NHR-49的核定位逐渐减少,就解释了为什么老年个体对“禁食大法”会没有响应。
归纳四点核心发现
这项研究把过氧化物酶体从“边缘角色”推到了舞台中央。它告诉我们:
1.过氧化物酶体的功能衰退是衰老相关的代谢僵硬的重要起始事件,而不是晚期结果。
2.核心蛋白PRX-5/PEX5的下降,导致细胞无法有效分解特定的多不饱和脂肪酸(如EPA),这些脂肪堆积、融合,变成难以动员的“顽固脂滴”。
3.过氧化物酶体失灵会依次“传染”给线粒体,导致其形态和能量代谢的双重崩溃。
4.饮食限制之所以能延寿,部分原因在于它维持了过氧化物酶体的功能;而直接过表达 prx-5 就足以改善脂滴和线粒体的衰老表型,并延长寿命。
当然,这项研究的主要模型是线虫,在人类中的转化还需要更多验证。
现阶段我们能做什么?
既然这项研究指出了过氧化物酶体的关键作用,那我们现在能不能做什么来“保护”它?还真有一些方向值得参考,但必须保持谨慎。
首先,饮食限制(即长期、适度的热量摄入控制)是目前证据最充分的干预手段。如果你能够科学地执行热量限制(例如在医生或营养师指导下减少每日总热量摄入,同时保证必需营养),确实有可能帮助维持细胞内的代谢协调。但请注意,这绝不等于“极端节食”,过度饥饿反而会导致肌肉流失和营养不良。

其次,不要自行补充高剂量EPA或相关鱼油。当过氧化物酶体功能失灵时,含EPA的甘油三酯会异常堆积,而不是被正常消耗(图3m)。盲目补充EPA不仅可能无效,反而可能加重脂质异常。如果你已经在服用鱼油补充剂,建议咨询医生进行个体化评估。
最后,保持规律的有氧运动。虽然研究中没有直接测试运动,但已知运动时,骨骼肌中高表达的PPAR-α会被激活,参与脂肪酸氧化的调控[4]。这可能是目前最安全的、辅助维持代谢灵活性的方式。
强调一下,真正的“过氧化物酶体修复药物”还处于基础研究阶段。在开发出能够稳定PEX5蛋白的小分子化合物之前,管住嘴、迈开腿,依然是高性价比的延缓衰老策略。
参考资料:
[1] Sharma A, Prabhakar A, Valera-Alberni M, et al. Peroxisomes orchestrate metabolic flexibility and longevity via an interorganelle cascade. Nature Aging. 2026;6(5):987-1006. doi:10.1038/s43587-026-01122-1
[2] Goodpaster BH, Sparks LM. Metabolic flexibility in health and disease. Cell Metabolism. 2017;25(5):1027-1036. doi:10.1016/j.cmet.2017.04.015
[3] Van Veldhoven PP. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. Journal of Lipid Research. 2010;51(10):2863-2895. doi:10.1194/jlr.r005959
[4] Manickam R, Duszka K, Wahli W. PPARs and microbiota in skeletal muscle health and wasting. International Journal of Molecular Sciences. 2020;21(21):8056. doi:10.3390/ijms21218056
