如果你关注过衰老领域的研究,大概率听说过“衰老细胞”和“衰老细胞清除药物”(senolytics,也叫希诺裂)这些概念。
简单来说,随着年龄增长,我们身体里会积累一批“老而不死”的细胞。它们不再分裂,也不干正事,却开始释放炎症因子,破坏周围组织。科学界已经公认,清除这些细胞可以延缓甚至逆转多种衰老相关的疾病,比如动脉硬化、骨关节炎、肺纤维化。
但有一个问题,一直没能完全解释清楚:为什么衰老细胞在老年组织里越积越多,而且赖着不“走”?
最近,《Nature Aging》发表了一篇观点文章,提出了一个很有意思的解释:问题可能不在细胞本身,而在它们“住”的地方——细胞外基质(EMC)[1]。
衰老细胞的两个反常行为
正常情况下,一个细胞如果受了重伤(比如DNA断裂、氧化应激),身体会启动程序让它“自杀”,也就是凋亡。但有些细胞受伤没那么严重,或者受伤的方式比较特殊,它们就会进入一种“休眠”状态,也就是细胞衰老。这是一种“半死不活”的状态:不再分裂,但也不会被清除,反而还要不停地往外“吐”各种有害分子——衰老相关分泌表型(SASP)。
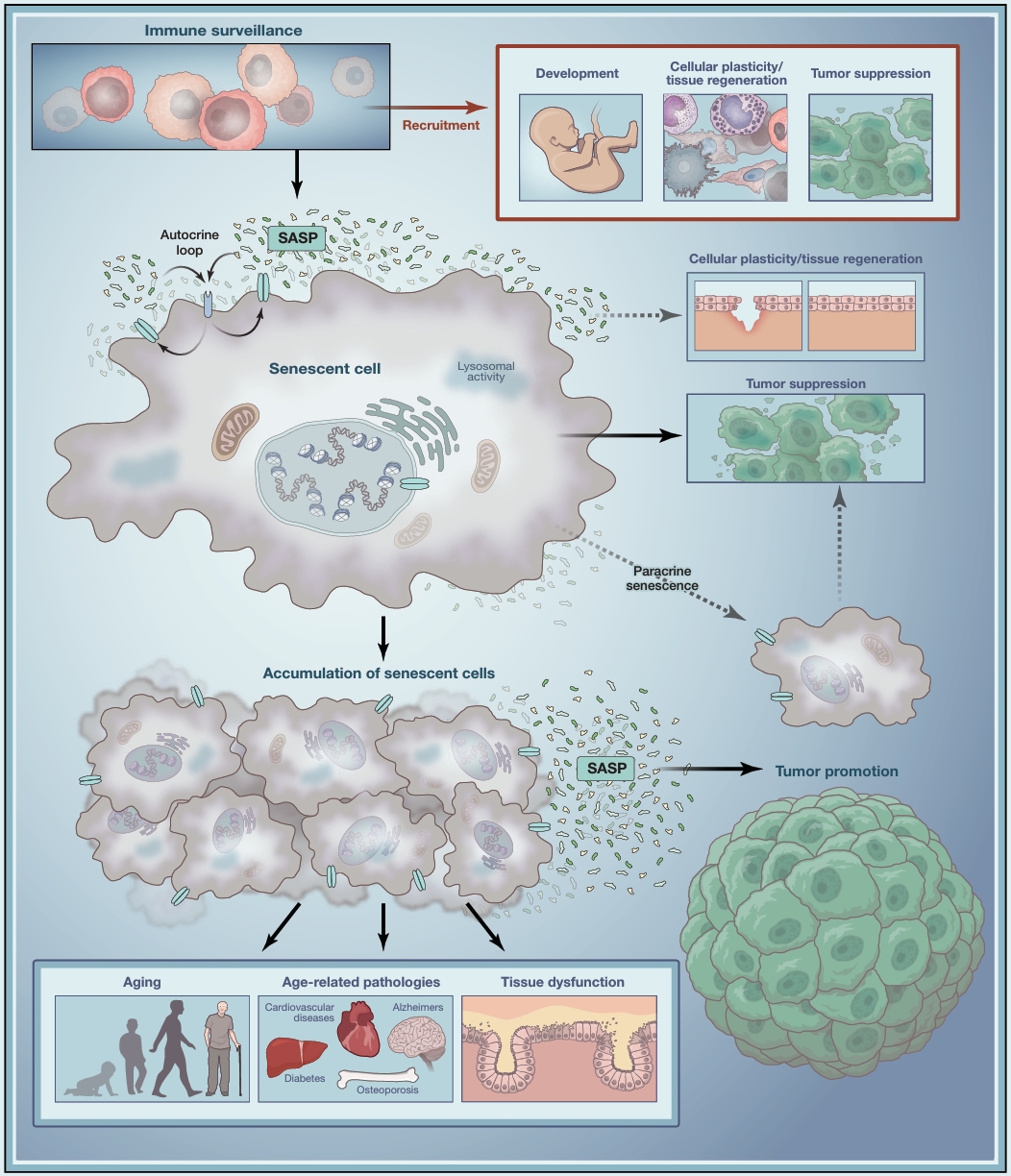
在年轻的健康组织里,这种衰老细胞通常会被免疫系统及时清除,不会造成大麻烦。但到了老年组织里,情况就变了。研究者观察到两个反常现象:
第一,同样程度的损伤,老年组织更容易“制造”出衰老细胞,而不是让它们凋亡。换句话说,细胞在老年环境里更容易选择“躺平”。
第二,一旦变成了衰老细胞,它们在老年组织里活得更久,不容易被清除。老年环境似乎在“保护”它们,好能让其长期存活。
这两个现象合在一起,就解释了为什么衰老细胞在老年组织中不断积累。那么问题来了:是什么在调控这两个环节?
传统的解释聚焦于细胞本身——比如DNA损伤累积、端粒变短、线粒体功能下降等。但这些“细胞内部”的理由,并不足以完全解释为什么衰老细胞的产生速度和存活时间会发生如此大的变化。于是,研究者把目光投向了细胞的外部环境。
细胞外基质:衰老进程中被忽视的主角
细胞不是孤零零地飘在液体里的,而是被一张巨大的“网络”包裹着,这张网络就叫细胞外基质。
你可以把它理解成细胞居住的“房子”。胶原蛋白、弹性蛋白、糖蛋白等各种纤维和填充物,就是房子的墙壁、地板、家具。但这栋房子并不是一成不变的,细胞外基质是一个动态的信号平台,它的成分、弹力、纤维的排列方式,都在向里面的细胞发送指令:“你该分裂了”“你该移动了”“你该分化了”……这些信号会通过细胞表面的受体(主要是整合素)传递到细胞内部。
细胞外基质的另一个重要功能是力学传导。细胞能够感知自己所处环境的物理特性,比如是坚硬还是有弹性。这种感知同样会通过整合素和细胞骨架来完成,然后转化成生物化学信号,告诉细胞“该做什么”。
然而,细胞外基质在衰老过程中会发生巨大的变化。多个物种、多个组织的研究都发现,随着年龄增长:
细胞外基质变得更坚硬(刚度增加);
胶原蛋白之间交联更多,弹性下降;
某些成分增多,另一些成分减少,整体“配方”发生改变。
这些变化不是被动的“老化”,而是主动影响细胞行为的信号。也就是说,房子本身变样了,住在里面的细胞收到的指令也就变了。
两个机制,解释两个谜题
说回这篇观点文章,它的核心贡献就是,把细胞外基质的变化与衰老细胞的两个反常行为直接联系了起来。具体来说,有两个机制。
机制1:变硬的细胞外基质让细胞更容易“选择衰老”
当细胞受到损伤时,它会面临一个“生死抉择”:是启动凋亡程序死掉,还是进入衰老状态躺平。研究者发现,整合素信号在这个抉择中扮演了关键角色。
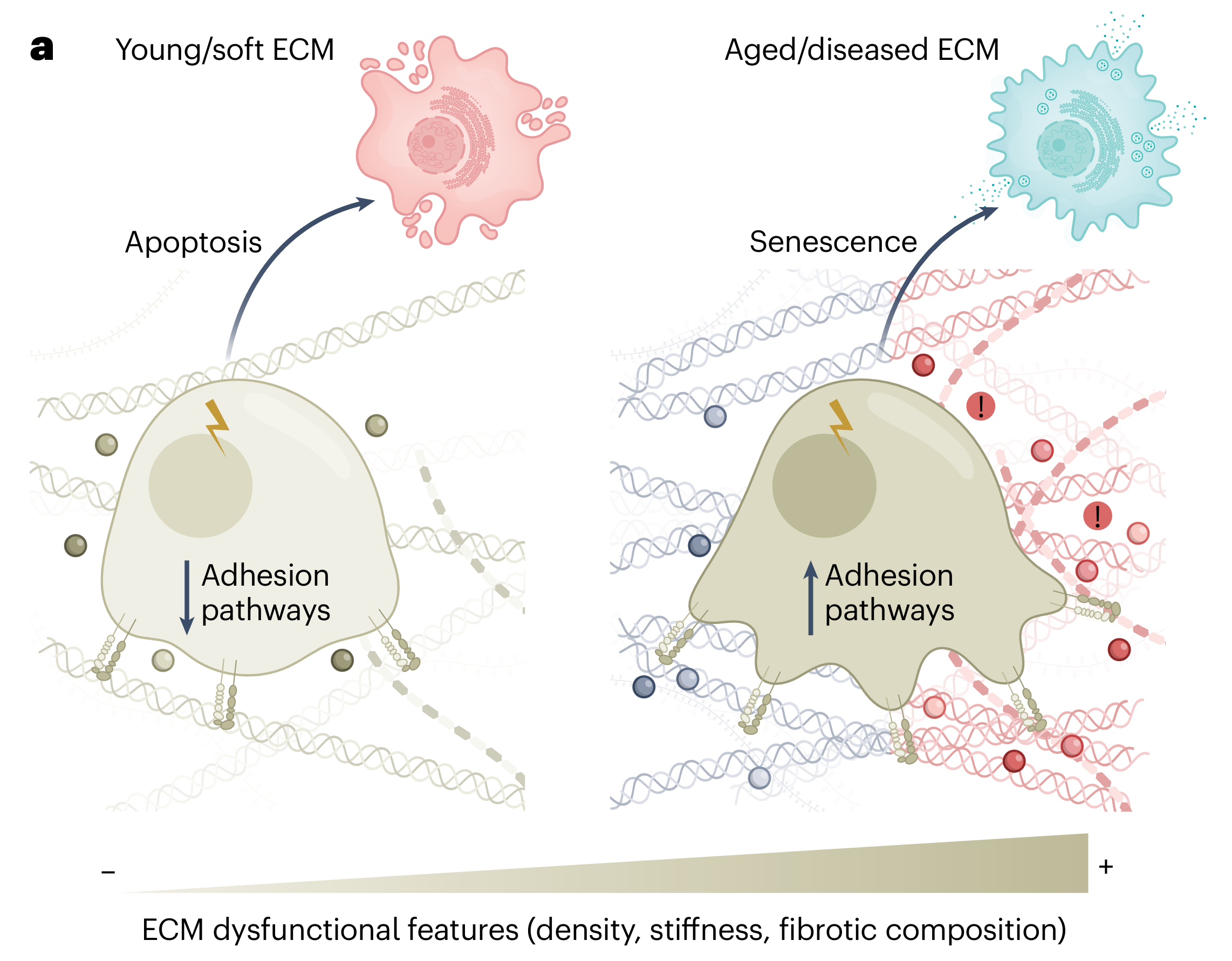
整合素是细胞表面的受体,专门感知细胞外基质的状况。当细胞外基质变硬、交联增多时,整合素会被更强烈地激活,进而启动下游一系列信号通路,其中一条重要的通路涉及一个叫做SRC的激酶(一种能磷酸化其他蛋白的酶)。
比如说,在亚致死性损伤条件下(即损伤程度不足以致死),如果整合素信号是完整的,SRC激酶被激活,细胞就会偏向于进入衰老状态。但如果用药物或基因手段阻断这条信号通路,同样的损伤就会让细胞转向凋亡。
换句话说,细胞外基质的物理状态直接改变了细胞对损伤的“解读”。同样的损伤,在柔软的基质环境里,细胞可能选择死亡;在坚硬、老化的基质环境里,细胞更可能选择衰老。
这就像同一个外伤,在干净的环境里伤口容易愈合,在脏乱的环境里容易感染——环境决定了结果。所以,老化变硬的细胞外基质,相当于把细胞的“生死开关”拨向了衰老那一侧。
机制2:变硬的细胞外基质让衰老细胞活得更久
衰老细胞一旦形成,也不是随随便便就能存活的。衰老细胞会大量分泌SASP,这种高强度的分泌活动,对细胞内部的内质网(负责合成和折叠蛋白质的细胞器)造成了巨大的压力。学术上叫“内质网应激”。
为了应对这种压力,衰老细胞需要依赖一条叫做YAP-TEAD的信号通路来维持内质网的稳定。YAP-TEAD是一个经典的“机械感知”通路,其活性高度依赖于细胞外基质的硬度。基质越硬,YAP-TEAD越活跃。
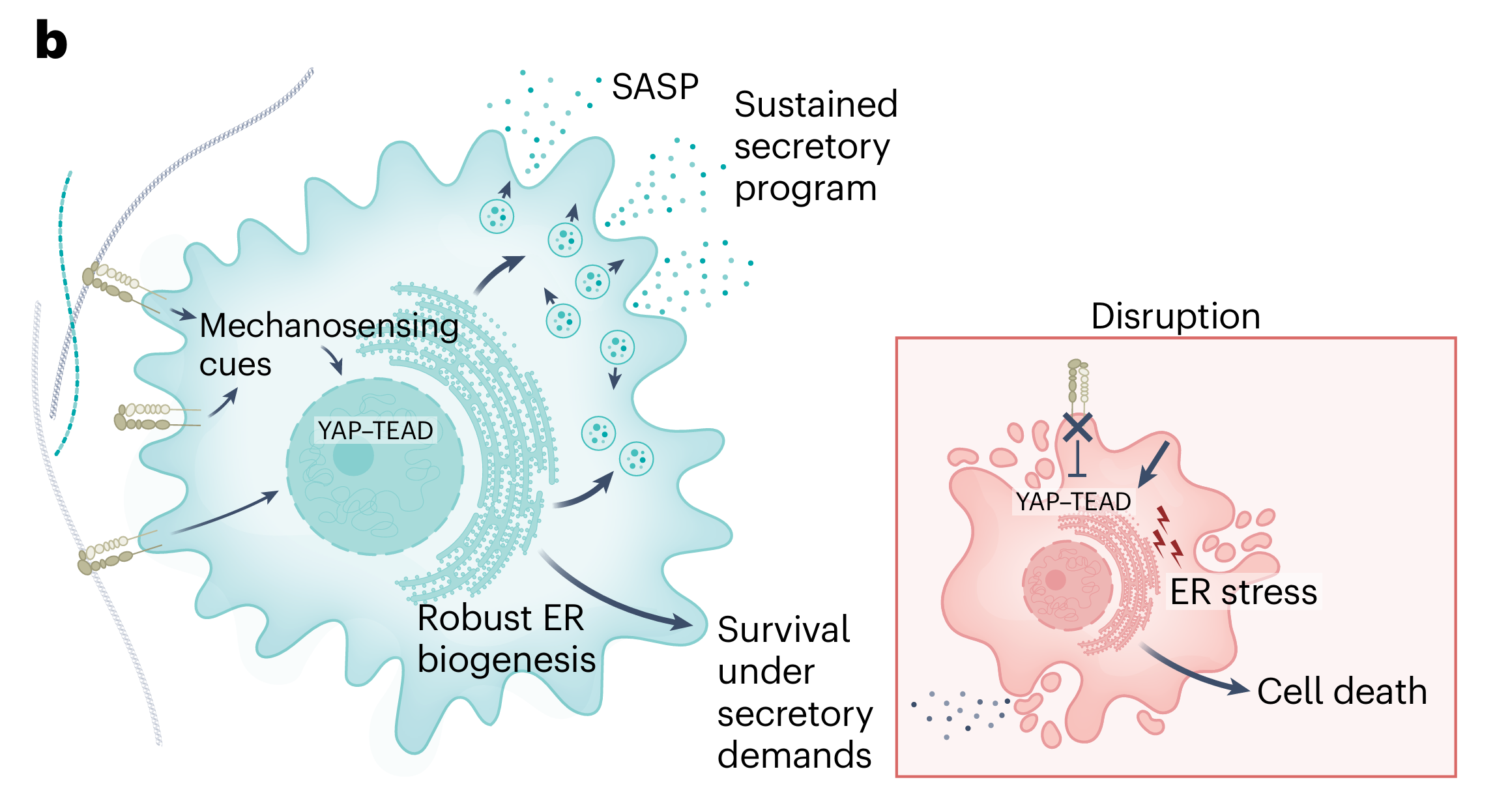
可以这样理解:衰老细胞像一台高负荷运转的机器,会产生大量热量(内质网应激),需要一套冷却系统(YAP-TEAD通路)来散热。而细胞外基质的硬度就是这套冷却系统的电源。在坚硬的老化基质里,电源稳定供电,冷却系统正常运转,机器可以一直转下去。如果把电源切断(抑制YAP-TEAD),机器就会过热烧毁。
目前广泛使用的衰老细胞清除药物“达沙替尼+槲皮素”中的达沙替尼,其部分作用机制就是抑制整合素下游的黏着斑激酶——整合素信号和力学传导的关键节点。也就是说,现有药物已经在无意中靶向了这个“房子信号”,只是我们之前没完全理解背后的逻辑。
恶性循环:衰老细胞反过来把房子搞得更糟
到这里,故事还没完。衰老细胞不只是被动地接受细胞外基质的信号,它们还会主动改造自己住的“房子”。
作为SASP的一部分,衰老细胞会分泌大量与细胞外基质重塑有关的分子:
某些胶原蛋白(让基质更密)
交联酶(比如赖氨酰氧化酶,让胶原纤维之间交联更严重,进一步增加硬度)
促纤维化因子(比如TGF-β,诱导周围细胞也产生更多基质)
结果就是:衰老细胞把自己的居住环境变得更硬、更密、更炎症,形成一个“更促进衰老”的环境。于是形成了一个正反馈循环:
老化/病变的细胞外基质 → 促进细胞进入衰老 → 衰老细胞改造基质 → 变得更老化的基质 → 进一步促进衰老。
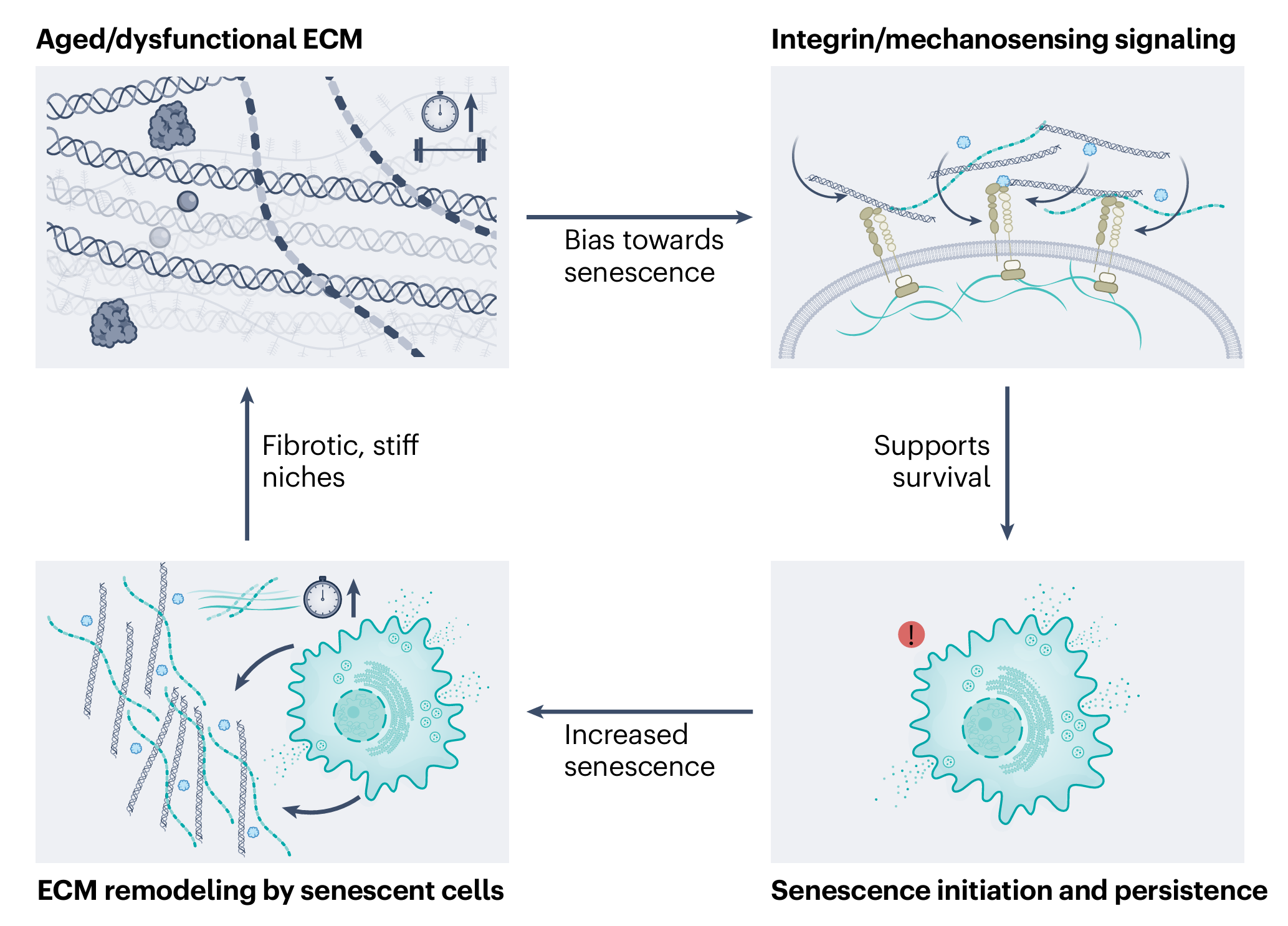
这个循环一旦启动,就会自我放大。这就解释了为什么在老年组织或慢性病变组织(如纤维化、慢性炎症)中,衰老细胞会异常富集,而且极难清除。
别只盯着衰老细胞,修复“房子”可能是更根本的策略
目前主流的抗衰老药物策略是,用希诺裂选择性地杀死衰老细胞。这个方法已经取得了不少进展,部分药物进入了临床试验。
但希诺裂有一个天然的局限:它只解决了“清除已有衰老细胞”的问题,并没有解决“新衰老细胞不断产生”的问题。只要老化细胞外基质这个“温床”还在,衰老细胞就会不断被制造出来,而且活得很好。
对此,文章提出了一个新的干预方向:把干预的重心从细胞本身,转移到细胞外基质和力学感知通路上来。具体来说,有两个潜在的干预路径:
路径一:靶向力学感知通路,破坏衰老细胞的生存依赖。比如使用YAP-TEAD抑制剂(维替泊芬已经在动物模型中验证可以减少衰老细胞),或者靶向上游的整合素信号节点,相当于切断衰老细胞的“保命电源”。
路径二:修复细胞外基质本身,让它恢复更“年轻”的状态。虽然听起来有点科幻,但其实已经有一些探索。例如,裸鼹鼠体内含有大量的高分子量透明质酸,这种分子可以改善细胞外基质的力学特性。研究者将裸鼹鼠的透明质酸合成酶转入普通小鼠,结果小鼠的健康寿命被延长,癌症发病率下降,总寿命也变长了。另外有研究表明,使用人工基质模拟年轻组织的成分和硬度,也可以逆转衰老细胞的某些功能异常。
这两条路径的核心思路是一样的:不要只追着衰老细胞打,要修复它们住的环境。环境修复好了,新衰老细胞自然减少,旧衰老细胞也更容易被清除。
普通人现在能做什么?
看到这里你可能想问:那我现在能做什么?先说清楚:这篇研究是基础科学框架,目前没有任何针对“ECM-机械感知回路”的疗法获批。别信任何声称能“重塑细胞外基质”的保健品。但从原理出发,有几件事确实值得做:
运动(最有效的ECM维护手段)
规律运动能直接改善ECM的弹性和成分,减少异常交联。中等强度的有氧运动(快走、慢跑、爬楼梯)加抗阻训练,每周150分钟。这是在帮细胞“修房子”。

控糖、减炎症
高血糖会加速胶原蛋白的糖基化交联(AGEs),是ECM硬化的重要原因。少吃精制糖和加工食品,同时管理好高血压、脂肪肝等慢性病,它们会诱发组织纤维化,加速ECM老化。
别浪费钱乱补胶原蛋白
口服胶原蛋白补剂会被消化成氨基酸,不一定补到你想补的地方。没有高质量证据证明它能逆转ECM的病理性老化。
在科研领域,一些新型衰老干预药物正在探索系统性调控炎症和代谢通路。例如,由赛诺根联合哈佛大学、梅奥诊所等机构研发的多靶点衰老干预候选药——SRN-901。在哺乳动物临床前试验中,接受SRN-901干预的小鼠中位剩余寿命延长33%,衰老速度减缓70%,显著下调了炎症反应、细胞凋亡等促衰老信号通路,并逆转了78种受衰老影响的代谢物。
现在能做的恰恰是那些“老生常谈”却靠谱的方法——运动、控糖、控制慢性炎症。这些措施的共同效果之一,就是延缓ECM的病理性老化,从而减少衰老细胞的产生和积累。
参考资料:
[1] Hernández-Bellido N, Perramon-Güell A, Anerillas C. The aging extracellular matrix as a missing link in senescent cell accumulation and persistence. Nature Aging. Published online May 4, 2026. doi:10.1038/s43587-026-01127-w
[2] Tian X, Azpurua J, Hine C, et al. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature. 2013;499(7458):346-349. doi:10.1038/nature12234
